| Tanulmányok : Szathmáry Eörs: Az élet keletkezése II. rész |
Szathmáry Eörs: Az élet keletkezése II. rész
2006.05.26. 20:08
A bioszintézis utak mintázatán alapuló csoportosítás érvényessége újabban kérdésessé vált, és csakugyan, a hipotézis erős formája nem tartható. Az "erős verzió" szerint az aminosavak hozzárendelődése a szomszédos kodonokhoz az előanyag-termék kapcsolat alapján valósult meg. Történetesen ez maga után vonta a kodonok és az aminosavak közötti koevolúciót: a kód pedig visszatükrözné az aminosavak megjelenését a bioszintézis során (Wong, 1975). Terres A. Ronneberg és munkatársai mostanában azt találták, hogy az előanyag-termék közti rokonsági kapcsolatokat régebben túl engedékenyen definiálták, és emiatt vált erősebbé a hipotézis. Tárgyilagosan szemlélve az erős változat láthatóan elpárolog.
6/b Az RNS-világtól az RNS-fehérje világig
Erősen korlátoznom kell magamat a RNS-világba ágyazott genetikai kód keletkezésének megvitatásában a következő megfontolások miatt: (1) bizonyítékaink vannak rá, hogy az RNS-világ anyagcseréje bonyolult lehetett; (2) az RNS-világból kivezető evolúciós út megvalósíthatónak látszik; (3) a most megoldásra váró probléma könnyebben bemutatható; (4) a hely hiánya megakadályoz abban, hogy más forgatókönyveket is tárgyaljak.
Majdnem közhely, hogy az RNS-világban keletkezett kezdetleges kódoló rendszer összes kritikus összetevőjének RNS természete kellett legyen. Feltételezhetően a ribozimek felismerték és módosították az aminosavakat (Szathmáry, 1990). Ez új fényt vet a genetikai kód hipotézisére, mely a régebbi sztereokémiai hipotézis egyfajta újjászületése, melynek értelmében a kódolás az aminosav és a kodon/antikodon között kialakuló közvetlen kapcsolaton alapul, hasonlóan a kulcs és a zár illeszkedéséhez. Az RNS-világ elmélete szerint egy ilyen kapcsolat be tud ágyazódni a nagyobb ribozimek alkotta anyagcserébe. Ez lenne a sztereokémiai felfogás erős formája, amely összefüggne az RNS-világgal. A gyenge változat azt mondaná, hogy a szintetáz ribozimek képesek az aminosavakhoz a kodon/antikodon hozzárendelést katalizálni, azonos módon, ahogyan ezt a mai fehérje szintetázok végzik. A szintetáz két különböző kötőhelyén felismeri az aminosavat és a megfelelő tRNS-t. Így ez indirekt sztereokémiai illeszkedés lenne a szintetáz ribozim RNS-motívuma és az aminosav között, ugyanakkor ez a kapcsolat sem a kodon, sem az antikodon részvételén nem alapulna (Szathmáry, 1993).
Michael Yarus (2000) kísérletei alapján azt javasolja, hogy az evolúció az RNS-világban az erős változattól a gyenge változat irányába mehetett. Úgy találta, hogy az aminosavak megkötésére mesterségesen szelektált RNS-molekulák aminosav kötőhelyein statisztikusan szignifikánsan gyakoribb a kodon (néha antikodon) szekvencia megjelenésének valószínűsége. Igaz ez az argininra, tirozinra és az izoleucinra, de nem igaz a fenilalaninra, habár az RNS képes az utóbbit is felismerni (Illangasekare - Yarus, 2002). Ezért a fenilalanin hozzárendelés során a gyenge változat érvényesülhetett.
Mindezek nyitva hagyják a kérdést, hogy mi az aminosav-kötés és -használat szelektív előnye az RNS-világban. Valószínűleg tartható az a javaslat, hogy az aminosavak (Szathmáry, 1993) vagy oligopeptidek (kevés aminosavból álló kovalens vegyületek) előnyösen összekapcsolódhattak az RNS-sekkel úgy, hogy a ribozimek katalitikus hatékonysága növekedjék (további részletek: Szathmáry, 1999). A hipotézis értelmében szintetáz ribozimek kicsi, tRNS-szerű molekulákhoz kapcsolták az aminosavakat, az anyagcsere ribozimei pedig ezeket az RNS "fogantyúkat" ragadták meg, amikor az aminosakra mint koenzimekre volt szükségük. A rövid RNS hajtűk csak a megfelelő aminosavhoz kapcsolódhattak (ez a "kódoló koenzim fogantyú" elképzelés: Szathmáry, 1993). A kódolás így előnyös lehetett fehérjeszintézis nélkül is, ami helyes magyarázata lehet ennek a nehéz evolúciós átmenetnek.
Idővel aztán több és több ilyen koenzim kapcsolódott a ribozimek szomszédos, egymást követő pozícióihoz (a tripletekhez). Így a szomszédos aminosavak még kötődtek a fogantyújukhoz, de ugyanakkor kiegészíthették egymás működését. Az aminosavak közül sokan az ősi peptidil-transzferáz ribozim - a mai egyik riboszomális RNS ősének - segítségével kapcsolódhattak össze egymással. A polipeptidek így méretben növekedhettek, míg a legtöbb ribozim hírvivő RNS-sé alakulhatott át.
Ez a forgatókönyv tisztán hipotetikus, de tesztelhető. Biztató, hogy Adam Roth és Ronald R. Breaker kísérlete szerint egy nukleinsav enzim hatékonyságát a sokszorosára lehetett növelni hisztidin kofaktor alkalmazásával. Nagyon sok kísérletre lesz még szükség, amíg a transzláció keletkezésének bonyolult problémáját kielégítően megoldjuk.
7. Kombinált megközelítési mód: egy kísérletes elősejt (őssejt) felé
7/a Mikor léptek színpadra az ősi sejtek?
Hasznos kérdés, hogy a chemoton logikai felépítése segíthet-e az életkeletkezés történeti folyamatának megértésében - mint azt az 1/d részben körvonalaztuk. Gánti lényegében empedoklészi álláspontra helyezkedett: elképzelése szerint a chemoton lehetséges alrendszerei egymástól függetlenül keletkeztek, majd az alrendszerek különféle kombinációi között szelekció mehetett végbe. Ez könnyen lehet, hogy téves megközelítés. Bár ígéretes próbálkozások történtek chemotonszerű rendszerek létrehozására, a nem enzimatikus anyagcsere sikeres létrehozása talán eleve lehetetlen feladat. Ezzel szemben számos vizsgálat alátámasztja azt a nézetet, hogy a sejt előtti evolúció egészen a ribozimek megjelenéséig folyhatott, jóval a chemotonszerű rendszerek spontán megjelenése előtt (vö. Maynard Smith - Szathmáry, 1995). Ha ez a nézőpont valóban helytálló, akkor a legtöbb kezdetleges, de valamelyest életképes chemoton már a kezdetektől ribozimeket tartalmazhatott.
Pier Luigi Luisi és munkatársai kísérletesen igazolták, hogy templát polimerizáció folyhat szaporodó lipid vezikulákon belül. Van két bökkenő. Először is, a templát polimerizációt egy kívülről adott fehérje enzim végezte. Másodszor, a templát polimerizáció nem volt összhangban a vezikula növekedésével és kettéosztódásával. Az anyagcsereciklus létrehozása szinte áthidalhatatlanul nagy probléma, mert a templát polimerizáció építőkövekkel való ellátása enzimek nélkül nem biztosítható megfelelő tisztaságban (l.: a Bevezetőt).
7/b Elősejtek anyagcsererendszer nélkül: a legszélsőségesebb heterotrófok?
Legújabban Jack W. Szostak és munkatársai (2001) körvonalaztak egy kísérleti programot, melynek célja az élet létrehozása egyszerű sejt formájában. Az elképzelt rendszer egy autokatalitikusan növekedő és kettéosztódó membránból és két specifikus ribozimből áll. Az első ribozim egy általános replikáz, amely képes saját maga, valamint a másik ribozim másolatainak előállítására. Utóbbi a lipid szintézisben vesz részt. Így a templát és a membrán dinamikája katalitikusan kapcsolt. A komplex előanyagokat, mint például az RNS replikáció monomerjeit, valamint a lipid prekurzorokat a vezikula készen venné fel a közegből. Számos kísérlet egyértelműen ebbe az irányba halad.
Lehetségesnek tűnik ilyesféle működő rendszert építeni, bár a membránon keresztüli anyagszállítás, valamint a templát replikáció és a membránnövekedés összehangolása további speciális problémákat vet fel (emlékezzünk, hogy a chemotonban e két alrendszer sztöchiometrikusan volt összekapcsolva).
8. Következtetések
Az élő rendszerek definíciójának munkahipotézise nélkülözhetetlen az életeredet elméleteinek megalkotásában és értékelésében. Az élő rendszer nem szükségszerűen evolúciós egység, és fordítva, az élőtől különböző egységek populációja is keresztülmehet a természetes szelekció által hajtott evolúción. De mivel bennünket elsősorban bioszféránk keletkezése érdekel, az első figyelemreméltó élőlény szükségképpen evolúciós egység is kellett hogy legyen. A legegyszerűbb élet egy hasznos és világos modellje a chemoton, mely három autokatalitikus rendszert tartalmaz: az anyagcsere-, a genetikai és a határoló alrendszereket. A rendszer mint egész képes a térbeli szaporodásra. Változatos elméleteket mutattunk be ezeknek az alrendszereknek a keletkezéséről és különböző összekapcsolásaikról. A kémiai evolúció területén végzett számos sikeres kísérlet ellenére még nem teljesen értjük a nukleinsavak és lipidek keletkezésének problémáját (ezek ma fontos összetevői az örökítő és határoló rendszereknek). E hiányosságok ellenére sokan hisznek abban, hogy az evolúciónak voltak átmeneti állapotai, mint például az RNS-világ, ahol az RNS-ek mint gének és mint enzimek funkcionáltak, ámbár prebiológiai szempontból elfogadható nem enzimatikus RNS önreplikációs mechanizmus még nem állt rendelkezésükre. A szerkezeti megfontolásokon kívül e rendszerek dinamikája is lényeges. Valószínű, hogy a kompartmentalizáció első passzív szakaszát (adszorpció az ásványi felszínhez) követte az aktív fázis (membrán általi elhatárolás), ami a kezdetleges genom felépülése szempontjából is elengedhetetlen lehetett. A genetikai kód valószínűleg egy olyan rendszerben születhetett, melyet már élőnek tekinthetünk. Az aminosavak a ribozimek koenzimjeiként kerülhettek az RNS-világba.
Kulcsszavak: élet keletkezése, replikátor, kémiai evolúció, autokatalis, templát replikáció, membrán, evolúció, RNS-világ, hibaküszöb, felületi anyagcsere
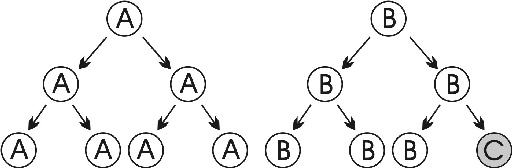 1. ábra * Az evolúció egységei. C egy új variáns. 1. ábra * Az evolúció egységei. C egy új variáns.
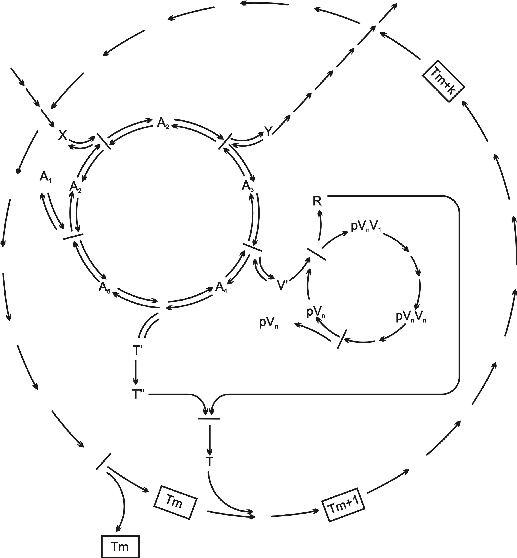 2. ábra * A chemoton modell absztrakt kémiai hálózata (Gánti után, 2003). Az Ai molekulák az autokatalitikus anyagcsereciklus köztitermékei (intermedierjei), mely előanyagokat (V') termel a templátreplikációhoz és a membrá növekedéshez (T'). A templát molekula pVn, n darab V molekulából áll. A membrán Tm, m darab T membránalkotó molekulából áll. Jól meghatározott feltételek mellet ez a rendszer képes növekedni és szaporodni a nyersanyag (táplálék, X) és hulladék (végtermék, Y) közötti energia- és anyagmennyiség-különbség terhére. 2. ábra * A chemoton modell absztrakt kémiai hálózata (Gánti után, 2003). Az Ai molekulák az autokatalitikus anyagcsereciklus köztitermékei (intermedierjei), mely előanyagokat (V') termel a templátreplikációhoz és a membrá növekedéshez (T'). A templát molekula pVn, n darab V molekulából áll. A membrán Tm, m darab T membránalkotó molekulából áll. Jól meghatározott feltételek mellet ez a rendszer képes növekedni és szaporodni a nyersanyag (táplálék, X) és hulladék (végtermék, Y) közötti energia- és anyagmennyiség-különbség terhére.
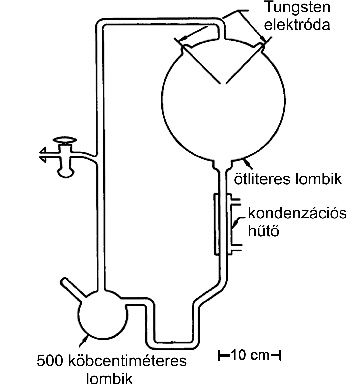 3. ábra * A klasszikus berendezés az "őslevesben" folyó kémiai evolúció vizsgálatához (Miller, 1998). A lombik a tetején redukáló gázokat, alul vizet tartalmaz. Elektromos kisülést a Tungsten elektródákkal állítanak elő. 3. ábra * A klasszikus berendezés az "őslevesben" folyó kémiai evolúció vizsgálatához (Miller, 1998). A lombik a tetején redukáló gázokat, alul vizet tartalmaz. Elektromos kisülést a Tungsten elektródákkal állítanak elő.
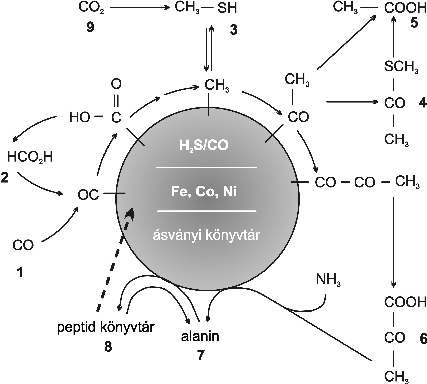 4. ábra * A kísérletek bizonyították, hogy magas hőmérsékleten és nagy nyomáson, a vas-kén világ forgatókönyve (Wächtershäuser, 2000) reális lehet. További információ a 3. táblázatban. 4. ábra * A kísérletek bizonyították, hogy magas hőmérsékleten és nagy nyomáson, a vas-kén világ forgatókönyve (Wächtershäuser, 2000) reális lehet. További információ a 3. táblázatban.
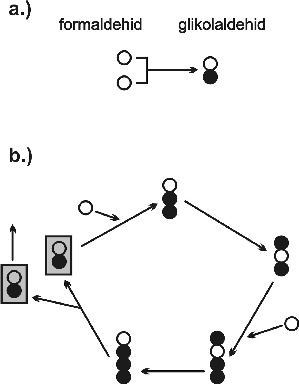 5. ábra * A formóz reakció, amely valójában az autokatalitikus cukorképzés komplex hálózata. (a) Az autokatalitikus mag "spontán képződése" nagyon lassú folyamat. (b) A hálózat autokatalitikus magja. Minden kör egy szénatomot tartalmazó csoportot jelent. 5. ábra * A formóz reakció, amely valójában az autokatalitikus cukorképzés komplex hálózata. (a) Az autokatalitikus mag "spontán képződése" nagyon lassú folyamat. (b) A hálózat autokatalitikus magja. Minden kör egy szénatomot tartalmazó csoportot jelent.
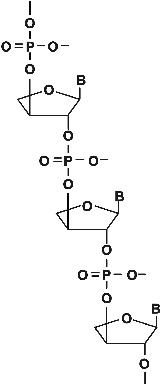 6. ábra * A treóz nukleinsav (TNA) az RNS lehetséges korai analógja (Joyce-tól, 2002). 6. ábra * A treóz nukleinsav (TNA) az RNS lehetséges korai analógja (Joyce-tól, 2002).
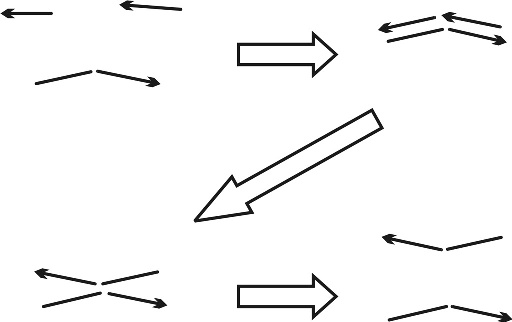 7. ábra * Majdnem exponenciálisan növekedő mesterséges replikátorok. A kialakuló termék molekuláris "gimnasztikája" csökkenti a termék általi gátlást: a komplex könnyebben esik szét, mint más esetekben. Meglepő, hogy ez megvalósítható peptid replikátorokkal is. 7. ábra * Majdnem exponenciálisan növekedő mesterséges replikátorok. A kialakuló termék molekuláris "gimnasztikája" csökkenti a termék általi gátlást: a komplex könnyebben esik szét, mint más esetekben. Meglepő, hogy ez megvalósítható peptid replikátorokkal is.
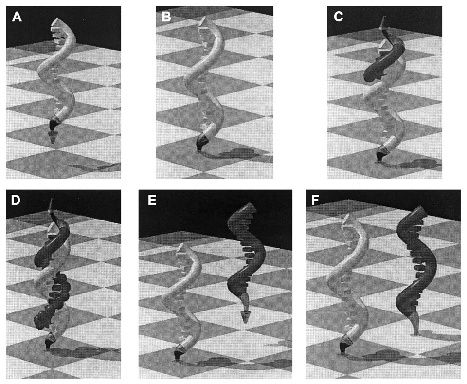 8. ábra * Von Kiedrowski SPREAD eljárása: a templátok exponenciális gyarapítása a felszín segítségével (Luther és mstai, 1998). (a) a templát (sárga) a reakció végéről származik. (b) a templát lehorgonyoz a felszínen. (c) a másolat (piros) szintézise: a lehorgonyzó vége a felszínnel ellentétesen van. (d) a másolódás befejezése. (e) a szálak szétválása után a másolat kész arra, hogy a felszínhez kapcsolódjon. (f) a templát és a másolat felszín segítségével választódik el egymástól, így mindketten új replikációba kezdhetnek. 8. ábra * Von Kiedrowski SPREAD eljárása: a templátok exponenciális gyarapítása a felszín segítségével (Luther és mstai, 1998). (a) a templát (sárga) a reakció végéről származik. (b) a templát lehorgonyoz a felszínen. (c) a másolat (piros) szintézise: a lehorgonyzó vége a felszínnel ellentétesen van. (d) a másolódás befejezése. (e) a szálak szétválása után a másolat kész arra, hogy a felszínhez kapcsolódjon. (f) a templát és a másolat felszín segítségével választódik el egymástól, így mindketten új replikációba kezdhetnek.
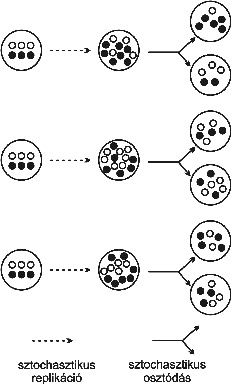 9. ábra * A szochasztikus korrektor modell. Az üres és fekete karikák a feltételezett őssejt kétféle génjét jelentik, a sejten belüli eltérő replikációsebességgel. Feltételezzük, hogy amikor a protosejt elkezdi a az életciklusát, a 3:3 templát összetétel az optimális az anyagcseréhez. Figyeljük meg, a két kompartment (a vastagon körbevettek) újból megjelenik, a belső versengés ellenére. 9. ábra * A szochasztikus korrektor modell. Az üres és fekete karikák a feltételezett őssejt kétféle génjét jelentik, a sejten belüli eltérő replikációsebességgel. Feltételezzük, hogy amikor a protosejt elkezdi a az életciklusát, a 3:3 templát összetétel az optimális az anyagcseréhez. Figyeljük meg, a két kompartment (a vastagon körbevettek) újból megjelenik, a belső versengés ellenére.
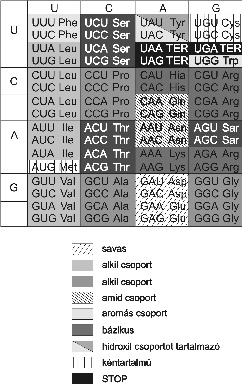 10. ábra * A kanonikus genetikai kód (Knight és mstai, 1999). Figyeljük meg a középső bázis domináns befolyását a rokon aminosavak kémiai természetére. Az ismert eltérések e genetikai "nyelv dialektusainak" tekinthetők. 10. ábra * A kanonikus genetikai kód (Knight és mstai, 1999). Figyeljük meg a középső bázis domináns befolyását a rokon aminosavak kémiai természetére. Az ismert eltérések e genetikai "nyelv dialektusainak" tekinthetők.
Baktériumok Emlősök Csillagközi jég Az üstökösök illó frakciója
Hidrogén 63,1 61,6 55 56
Oxigén 29,0 26,0 30 31
Szén 6,4 10,5 13 10
Nitrogén 1,4 2,4 1 2,7
Kén 0,06 0,13 0.8 0,3
Foszfor 0,12 0,13 - (0,08)
Kalcium - 0,23 - -
1. táblázat * Az elemek gyakorisága az élő szervezetekben és az űrben. Az adatok százalékban vannak megadva. Az üstökösök foszfortartalmát az űrben található sűrűségből számolták. Delsemme (1998) könyvéből, p. 114.
arginin porfirinek riboflavin
lizin piridoxál folsav
hisztidin tiamin liponsav
egyenes szénláncú zsírsavak biotin
2. táblázat * Prebiotikus körülmények között sikeresen szintetizált, biológiai fontosságú vegyületek. Miller (1998) nyomán.
Reakció Katalizátor Hőmérséklet Nyomás
(1) -> (2) (Fe,Ni)S 100C 0,2 Mpa
(1) -> (3) (Fe,Ni)S 100C 0,2 Mpa
(9) -> (3) FeS 100C 0,2 Mpa
(1) -> (5) (Fe,Ni)S 100C 0,2 Mpa
(3) -> (4) (Fe,Ni)S 100C 0,2 Mpa
(2) -> (6) FeS 250C 200 Mpa
(6) -> (7) FeS 100C 0,2 Mpa
(7) -> (8) (Fe,Ni)S 100C 0,2 Mpa
3. táblázat * A 4. ábrán bemutatott hálózat folyamatainak feltételei. Wächtershäuser (2000) nyomán.
Kialakított kötésb Távozó csoport A ribozim aktivitása
-O-PO3- 5'-RNA Foszfodiészter hasításc
HO-PO3- Ciklikus foszfát hidrolízisec
-O-PO3- PPi RNS ligáció
-O-PO3- PPi Korlátos polimerizáció
-O-PO3- AMP RNS ligáció
-O-PO3- ADP RNS foszforiláció
-O-PO3- Imidizole Tetrafoszfát sapka képzése
-O-PO3- Rppd Foszfát anhidrid transzfer és hidrolízis
-O-PO3< PPi RNS elágaztatása
-O-CO- AMP RNA aminoacilálása
-O-CO- 3'-RNA Acil transzfer
-O-CO- AMP Acil transzfer
-HN-CO- 3'-RNA Amid kötés kialakítása
-HN-CO- AMP Peptid kötés kialakítása
>N-CH2- I RNS alkiláció
-S-CH2- Br Thio alkylation
>HC-CH< Diels-Alder addíció (antracén-maleimid)
>N-CH< PPi Glikozidos kötés képzése
Hidas bifenil izomerizációja
Porfirin metiláció
a Ezeket az aktivitásokat DNS és egyéb RNS-analógok készletéből is izolálták.
b A támadó nukleofil balra van. A kialakított kötést (-) jelöli.
A szomszédos, egy (-) vagy kettő (< vagy >) atomhoz vezető kötéseket jelöltük.
c A katalízis az RNS újfajta feltekeredésének köszönhető.
d Az R nagyszámú különböző csoport bármelyike lehet.
4. táblázat * Néhány újonnan in vitro szelektált ribozima, Bartel és Unrau nyomán (1999).
IRODALOM
Czárán Tamás - Szathmáry Eörs (2000): Coexistence of Replicators in Prebiotic Evolution. in Dieckmann, Ulf - Law, Richard - Metz, Johan A. J. (eds.): The Geometry of Ecological Interactions: Simplifying Spatial Complexity. IIASA - Cambridge University Press, 116-134.
Crick, Francis H. C. (1968): The Origin of the Genetic Code. Journal of Molecular Biology. 38, 367-379.
Crick, Francis H. C. - Brenner, S. - Klug, A. - Pieczenik, G. (1976): A Speculation on the Origin of Protein Synthesis. Origins of Life. 7, 389-397.
Eigen, Manfred (1971): Self-Organization of Matter and the Evolution of Biological Macromolecules. Naturwissenschaften. 58, 465-523.
Gánti Tibor (1971): Az élet princípiuma. Gondolat, Bp.
Gánti Tibor (2003): The Principles of Life. Oxford Univ. Press, Oxford
Illangasekare, Mali - Yarus, Michael (2002): Phenylalanine-Binding Rnas and Genetic Code Evolution. Journal of Molecular Evolution. 54, 298-311.
Kiedrowski, Günter von (1986): A Self-Replicating Hexadeoxy Nucleotide. Angewandte Chemie International Edition in English. 25, 932-935.
Kiedrowski, Günter von (1993): Minimal Replicator Theory I: Parabolic Versus Exponential Growth. Bioorganic Chemistry Frontiers. 3, 113-146.
Kiedrowski, Günter von (1999): Molekulare Prinzipien Der Artifiziellen Selbstreplikation. in Ganten, Detlev (ed.): Gene, Neurone, Qubits & Co. Unsere Welten Der Information. S. Hirzel Verlag, Stuttgart, 123-145.
Kiedrowski, Günter von - Szathmáry Eörs (2000): Selection Versus Coexistence of Parabolic Replicat-ors Spreading on Surfaces. Selection. 1, 173-179.
Knight, Robin D. - Freeland, Stephen J. - Landweber, Laura F. (1999): Selection, History and Chemistry: Three Faces of the Genetic Code. Trends in Biochemical Sciences. 24, 241-247.
Luisi, Pier Luigi (1998): About Various Definitions of Life. Origins of Life and Evolution of the Biosphere. 28, 613-622.
Luther, Anatol - Brandsch, Romana - Kiedrowski, Günter von (1998): Surface-Promoted Replication and Exponential Amplification of DNA Analogues. Nature. 396, 245-248.
Maynard Smith, John (1979): Hypercycles and the Origin of Life. Nature 280, 445-446.
Maynard Smith, John (1986): The Problems of Biology, Oxford University Press, Oxford
Maynard Smith - Szathmáry (1995): The Major Transitions in Evolution. Freeman, Oxford
Maynard Smith, John - Szathmáry Eörs (1999): The Origins of Life. Oxford University Press, Oxford
Oparin, Alexander Ivanovich (1961): Life: Its Nature, Origin and Development. Academic Press, N.Y.
Orgel, Leslie E. (1968): Evolution of the Genetic Apparatus. Journal of Molecular Biology. 38, 381-393.
Orgel, Leslie E. (2000): Self-Organizing Biochemical Cycles. Proceedings of the National Academy of Sciences of the USA. 97, 12503-12507.
Rebek, Julius (1994): Synthetic Self-Replicating Molecules. Scientific American. 271, 1, 34-40.
Segré, Daniel - Ben-Eli, D. - Deamer, D. W. - Lancet, D. (2001): The Lipid World. Origins of Life and Evolution of the Biosphere. 31, 119-145.
Szabó Péter - Scheuring I. - Czárán T. - Szathmáry E. (2002): In Silico Simulations Reveal that Replicators with Limited Dispersal Evolve towards Higher Efficiency and Fidelity. Nature. 420, 360-363.
Szathmáry Eörs (1990): Towards the Evolution of Ribozymes. Nature. 344, 115.
Szathmáry Eörs (1991): Simple Growth Laws and Selection Consequences. Trends in Ecology and Evolution. 6, 366-370.
Szathmáry Eörs (1993): Coding Coenzyme Handles: A Hypothesis for the Origin of the Genetic Code. Proceedings of the National Academy of Scienes of the USA. 90, 9916-9920.
Szathmáry Eörs (1999): The Origin of the Genetic Code. Amino Acids as Cofactors in an RNA World. Trends in Genetics. 15, 223-229.
Szathmáry Eörs (2000): The Evolution of Replicators. Philosophical Transactions of the Royal Society of London B. 355, 1669-1676.
Szathmáry Eörs (2002): Units of Evolution and Units of Life. in Pályi Gyula - Zucchi, Claudia - Caglioti, Luciano (eds.) Fundamentals of Life. Elsevier, Paris, 181-195.
Szathmáry Eörs - Demeter László (1987): Group Selection of Early Replicators and the Origin of Life. Journal of Theoretical Biology. 128, 463-486.
Szathmáry Eörs - Gladkih, I. (1989): Sub-Exponential Growth and Coexistence of Non-Enzymatically Replicating Templates. Journal of Theoretical Biology. 138, 55-58.
Szathmáry Eörs - Maynard Smith, John (1995): The Major Evolutionary Transitions. Nature. 374, 227-232.
Szathmáry Eörs - Maynard Smith, John (1997): From Replicators To Reproducers: The First Major Transitions Leading To Life. Journal of Theoretical Biology. 187, 555-571.
Szathmáry Eörs - Zintzaras, Elias (1992) A Statistical Test of Hypotheses on the Organization and Origin of the Genetic Code. Journal of Molecular Evolution. 35, 185-189.
Szostak, Jack W. - Bartel, David P. - Luisi, Pier Luigi (2001): Synthesizing Life. Nature. 409, 387-390.
Wächtershäuser, Günter (1988): Before Enzymes and Templates: Theory of Surface Metabolism. Microbiological Reviews. 52, 452-484.
Wächtershäuser, Günter (1992): Groundworks For An Evolutionary Biochemistry: The Iron-Sulfur World. Progress in Biophysics and Molecular Biology. 58, 85-201.
Woese, Carl R. (1967): The Genetic Code. Harper & Row, New York
Wong, J. T.-F. (1975): A Coevolution Theory of the Genetic Code. Proceedings of the National Academy of Science of the U.S.A. 72, 1909-1912.
Yarus, Michael (2000): RNA-Ligand Chemistry: A Testable Source For the Genetic Code. RNA. 6, 475-484.
Zintzaras, Elias - Santos, Mauro - Szathmáry Eörs (2002): “Living" Under the Challenge of Information Decay: The Stochastic Corrector Model Versus Hypercycles. Journal of Theoretical Biology. 217, 167-181.
| 


